#19,202
Yesterday we looked at an H9 avian flu infection in a Hong Kong toddler who's only known risk exposure was visiting a live market. While not exactly a smoking gun, the CHP's epidemiological investigation stated `. . . It cannot be ruled out that the patient was infected through indirect contact with a contaminated environment at the wet market.'
The link between live bird markets and the spread (and potential reassortment of) avian flu strains has been long established. LBMs typically bring together birds of varying species (chickens, ducks, geese, quail, and others) - often imported from different farms - which are housed in cramped quarters.
Twelve years ago, in CDC: Risk Factors Involved With H7N9 Infection we looked at a case-control study conducted by an international group of scientists, including researchers from both the Chinese and the US CDC that concluded.
Exposures to poultry in markets were associated with A(H7N9) virus infection, even without poultry contact. China should consider permanently closing live poultry markets or aggressively pursuing control measures to prevent spread of this emerging pathogen.
In 2016's Interventions in live poultry markets for the control of avian influenza: A systematic review Vittoria Offeddu , Benjamin J. Cowling, and J.S. Malik Peiris laid out the risks of avian influenza from live bird markets, reviewed some of the possible interventions, and concluded:
Highlights
- Avian influenza viruses (AIVs) can infect humans. Bird-to-human transmission is particularly intense in live poultry markets.
- Periodic rest days, overnight depopulation or sale bans of certain species significantly reduce AIV-circulation in the markets.
- Market closure would lastingly reduce the risk of animal and human infection.
- keep live poultry in their backyards or homes, or who purchase live birds at markets;
- slaughter, de-feather and/or butcher poultry or other animals at home;
- handle and prepare raw poultry for further cooking and consumption;
Despite this tarnished reputation, and repeated calls to close them, LBMs flourish around the world.
While most common in Asia and the Middle East, live are also found in Europe and the United States. Last March, in USDA Report 9 More Live Bird Markets Infected With HPAI H5 we looked at outbreaks in 3 states (New York, Florida, Pennsylvania).Earlier this week the USDA reported a new outbreak at a live Market in Passaic, New Jersey.

Interestingly, Passaic was one of 3 wastewater monitoring sites in the U.S. that reported HPAI H5 positives in the latest CDC report.

All of which brings us to the second report this week, this time from Rhode Island, where a routine quarterly inspection found asymptomatic H5-positive poultry. This press release from the Rhode Island Department of Environmental Management.
Avian Flu Confirmed at Live Bird Market in Providence
Published on Saturday, June 13, 2026
The Rhode Island Department of Health (RIDOH) and Rhode Island Department of Environmental Management (DEM) want to alert consumers that birds at Antonelli Poultry in Providence tested positive for the H5N1 strain of avian influenza during routine quarterly testing by the US Department of Agriculture (USDA). The infected birds, which included live chickens and ducks, did not come from Rhode Island farms. They were from out-of-state dealers.
Earlier today (June 13), the State Veterinarian oversaw the USDA-required humane euthanasia of about 445 asymptomatic birds at the market to prevent the spread of the disease to other birds. Per USDA regulations, Antonelli Poultry will be closed until 5 days after they have disposed of infected birds and have cleaned and sanitized all areas of the business. Antonelli Poultry is closely cooperating with DEM and RIDOH.
Because staff at Antonelli Poultry may have been exposed to avian influenza, and out of an abundance of caution, RIDOH is monitoring all staff for 10 days for symptoms of avian influenza. The overall risk of humans getting H5N1 remains low.
“Cooking poultry to the proper internal temperature of 165° kills bacteria and viruses, including avian influenza A viruses,” says Director of Health Jerry Larkin, MD. “RIDOH recommends that if anyone still has poultry they bought between June 9 and June 12 that was killed and dressed by Antonelli Poultry, they should double bag the poultry and dispose of it in their regular trash. If you have properly cooked and eaten chicken from Antonelli Poultry, the risk of becoming ill is very low; however, if you develop symptoms of avian influenza, you should seek medical care.”
Symptoms of avian influenza include eye redness, fever, cough, sore throat, runny nose, muscle or body aches, fatigue, shortness of breath or difficulty breathing, or pneumonia that requires hospitalization. People who get avian influenza can be treated with antivirals.
To prevent any foodborne illness, RIDOH recommends:
Avian influenza infections in humans are rare. The best way to prevent avian influenza in humans is for people to avoid exposure.
- Wash hands, utensils, and cutting boards before and after contact with raw poultry, meat, seafood, and eggs.
- Keep raw poultry and meat away from food that won’t be cooked—like fruits and vegetables.
- Cook food to the proper temperature and use a food thermometer to check the food’s internal temperature. You cannot tell by looking at food if it is cooked to the proper temperature.
“DEM works closely with federal and State veterinary and public health officials to respond quickly to confirmed H5N1 cases in domestic birds,” said State Veterinarian Scott Marshall, DVM. “The USDA performs quarterly testing at live bird markets to ensure the public’s safety.”
- Avoid direct contact with birds or other animals infected with, or suspected to be infected with, avian influenza.
- Avoid direct contact with sick or dead wild birds, poultry, or other animals.
- Do not touch surfaces or materials contaminated with saliva, mucous, or animal feces from wild or domestic birds or other animals with confirmed or suspected avian influenza.
- Do not touch or drink raw milk (unpasteurized milk), especially from animals with confirmed or suspected avian influenza
- Do not handle any sick or dead wild birds or other animals without wearing personal protective equipment (PPE).
This is Rhode Island’s first confirmed domestic bird case of avian influenza in 2026. Rhode Island has previously confirmed infections in noncommercial flocks in 2022 and in 2025.
To learn more about avian influenza in humans, visit RIDOH’s website. To learn more about avian influenza in animals, visit DEM’s website.
Date Sat, 06/13/2026 - 16:02
Notably, the press release stated that the birds had been imported from another (unnamed) state. Worth noting, we've not seen any commercial flocks reported by the USDA as H5 positive east of Indiana since early May.Government agencies are quick to reassure the public that the risk of contracting avian influenza remains low in the United States, but exposure to live birds (via LBMs or raising poultry) is a known risk factor (see CDC graphic below).
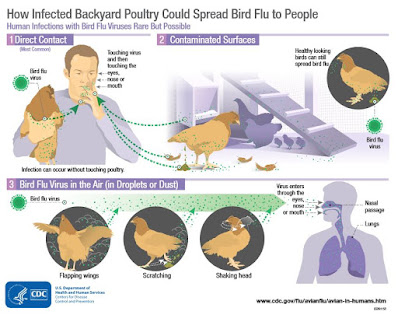
Over the past few years the threat from H5N1 has grown markedly in the Western Hemisphere. Things we used to do without much thought - like raising a few chickens the backyard or frequenting live markets - carry more risks today.
While those risks can be largely mitigated through improved biosecurity practices, four years after its arrival, we still seem to be tempting fate.